*
\\// Home \\//Genetics Theory \\// Chicken Mutations\\// Chromosome Linkages\\//
\\//Chicken Variety Genotypes\\//Genetics Calculators\\//
\\// References/Links\\// About / Contact Us\\//
Old URL: http://home.ezweb.com.au/~kazballea/genetics/mutations1.html
Last updated: 4 Jan., 2009
* Most of the photos on this website originated from "Feathersite",
and "Old English Bantam Club of America" websites, and have been given with
permission by the website administrators.
© Copyright of images belongs to cited photography contributors.
-----------------------------------------------------------------------------
* ![]() Click here for "Print Version"
Click here for "Print Version"
* ![]() Download (..rignt click_Save Target As..) "PDF Version (495 KB)"
Download (..rignt click_Save Target As..) "PDF Version (495 KB)"
-----------------------------------------------------------------------------
This section became quite large, so I've divided the mutation section into three pages:
Mutations: <<Page 1>><<Page2>><<Page3>>
|
Plumage Colour/Pattern genes
Feather mutations
Leg/Skin colour
Eye Colour & ear lobe colour
Physical Mutations
Size Mutations
|
Below are the E locus genes.
Genes: E, ER, ER- Fay, eWh, e+, eb, es, ebc, ey, eq
Gene names: Extended Black, Birchen, Fayoumi Birchen, Dominant Wheaten, wild type, Brown, Speckled, Buttercup, Recessive Wheaten, Queen SilviaPhenotypes: Black, Brown Red, Brown Red, Wheaten, Black breasted Red (salmon breasted hens), Brown (brown breasted hens), Speckled, Buttercup, Clay Wheaten, Queen Silvia.
* note: "ER- Fay" is not recognised nomenclature for the Fayoumi Birchen allele, only used here for short hand purposes. Smyth (1994) listed a Birchen allele as ERB, the RB standing for "red-brown downed birchen". Possibly this is the Birchen allele found in Fayoumi fowl.
.
Above are the typical e locus down patterns, without columbian-like restrictors (except maybe ebc chick- Db). Columbian (Co) will change the appearance of e+ & eb down (usually adds eumelanin pigment, particularly on the back), but not E, ER & eWh (although eWh with Co are changed to a buff colour if sex-linked gold is present, but remain cream if sex-linked silver). Both E & ER down are indistinguishable, with variation from all black (eumelanin) to black with white as shown above.
The E locus is important to learn with poultry genetics. The common E locus alleles are:
The E locus alleles produce the base primary colour/patterns to which varieties are build upon. The e+ allele is the wild -type, and produces the typical Black Breasted Red male- salmon breasted female phenotypes (as found in the Red Jungle Fowl, etc). All male phenotypes, except E & ER, have the Black Breasted Red colour/pattern.
* Images used in the following photos are from Feathersite, and the OEGBCA website.

* Wild type e+ adult plumage and chick down

* Wheaten eWh adult plumage and chick down

* Brown eb adult plumage and chick down

* Birchen ER adult plumage and chick down
-------------------------------------------------------
It is a recent occurrence the DNA sequencing of two Birchen alleles, as once previously thought of as only one allele - ie ER Birchen. The following is a direct quote from Dr Ron Okimoto on the Extended Black and two Birchen alleles (quote from Polish Chickens - A Discussion of E - extended black ):
Dr Ron Okimoto......." We have found three different alleles of the melanocortin 1 receptor locus (MC1-R) that produce black downed chicks on an otherwise wild-type background. Our data indicates that the MC1-R gene is the E locus. The most dominant allele at this locus is the E extended black allele. It differs from wild-type e+ by two mutations that change the amino acid sequence of the gene (Met71Thr and Glu92Lys). The Leghorn birchin allele we call ER only has the Glu92Lys mutation and lacks the Met71Thr. Our hypothesis is that you need both mutations to get the blackest bird and that if you have only the Glu92Lys you end up with the birchin black phenotype. There is a third ER allele that is found in Fayoumi. The Fayoumi birchin allele has a totally different mutation (Leu133Gln).
The most dominant E allele has varied expression in down color. Some chicks are black with gray bellies, and other show varying amount of cream color on their undersides. This color seems to be due to other genes than the E locus because I get both types of chicks with the most dominant E allele."
The following is another direct quote from Dr Okimito, first discussing the E and ER alleles, then eb & eWh (quote from The Classroom @ The Coop -Different Kinds of Black ):
Dr Ron Okimoto......"At the molecular level these alleles seem to be related. Birchin was probably the first mutation. It has the same mutation that causes black fur in some mouse lines. The E allele probably came second in addition to the birchin mutation the E allele has a second mutation at the same position that causes black wool in sheep. So the E allele has a double whamy that produces more black than the birchin mutation by itself can. The eb allele has a third mutation. This just means that there was a bird with the E allele and it mutated to the eb allele. This third mutation seems to be unique among animals and appears to act to attenuate the E allele so that the birds aren't black, but brown.
The wild-type e+ allele has none of these mutations. The dominant wheaten allele has a unique mutation that separates it from e+. I haven't found a recessive wheaten allele yet. All wheaten samples that I've tested look identical to dominant wheaten. This may mean that the guys that claimed that other modifiers determined the dominance of wheaten may be correct."
The following is direct quote from Dr Okimoto on the DNA sequencing of E locus allele from the Silician Buttercup breed, by Leif Anderson. This indicates that the Buttercup pattern may be produced with the eb allele in combination with other modifying genes, and that the ebc allele had not been sequenced.
Dr Ron Okimoto (quote from The Classroom @ The Coop) ......." Leif Andersen has sequenced the buttercup allele (ebc) and it is the identical sequence to the eb alleles that I've sequenced in Partridge Rocks and the Smyth Brown line. If there is a difference between the alleles it is in the regulation of the gene and not in the coding sequence. There may be linked genes that affect the expression of the E locus too."
Further research by Dr Okimoto (quote from Classroom @ The Coop: Blue & E/ or ER/, posted May 02, 2006) ......
Some of us speculated that Andalusian blue used ER instead of E because ER would be more amenable to secondary pattern genes like Pg.
I recently tested some Andalusian Blues from McMurray. I had a mixed batch of chicks so I could only tell the gray ones as blues the blacks could have been something else. There were three gray chicks that produced gray chick feathers. Two were homozygous ER ER and the third was heterozygous E ER.
My guess is that E probably causes the dark birds and that the show quality birds are probably ER.
I had ordered 5 birchin Cochin bantams, but I may have gotten black Cochins by mistake because all 5 feather footed black chicks were homozygous for E and not ER. I couldn't tell if there had been a mix up because we couldn't raise the chicks longer than two weeks in the isolation facilities. I found two weird alleles in McMurray Buff Rocks. I was looking for recessive wheaten, and it may be in those stocks. One allele has the dominant wheaten mutation, but it is coupled with the Fayoumi birchin mutation. You can speculate and infer that it might produce more black melanin than dominant wheaten.
There is another allele that has the eb attenuating His/Pro mutation that turns the extended black allele to a brown allele, but the attenuating mutation is in the ER sequence. My guess is that this allele produces less black than the eb allele, how much less is anyones guess. Someone would have to cross these Buff birds out to Brown Leghorns and backcross them to Leghorns while tracking the E locus alleles and getting rid of the columbian restrictors.
Dr Ron Okimoto on E locus alleles order of dominance, and Recessive Wheaten..... (quote from E-locus: Order of dominance (again!) , posted August 12, 2006) ......
Even E isn't completely dominant on some genetic backgrounds. In some cases I've gotten eWH ER heterozygotes that look like eb birds.
Recessive wheaten is a strange story, but Smyth has told me that he used to maintain a recessive wheaten line, and Morejohn had a Red Junglefowl line that segregated wheaten downed chicks from wild-type parents in 3:1.
There probably is a recessive wheaten somewhere. I found a Buff Rock line that segregates for a weird E locus allele that seems to be a double mutant. I got it from McMurray. It has the Fayoumi birchin mutation and the dominant wheaten mutation in the same gene. I don't know what the phenotype of such a bird would be like. The Fayoumi mutation would send signal all the time (produce black pigment) and the wheaten mutation would inhibit signal propagation. At least, that is the theory if wheaten is like red fur color in mammals.
The idea is that dominant wheaten is dominant because of haplo insufficiency. This just means that in the presence of wheaten not enough signal is sent by the e+ or eb allele product and you get wheaten instead of stippled. Recessive wheaten would produce enough signal so that heterozygotes could still produce stippling, but homozygous recessive wheatens could not because two low signal producing gene products could not produce enough signal to produce stippling.
You can look at it like this, say that eWh eWh homozygotes produced 0 signal, eWh e+ heterozygotes produced 1/2 signal, but it isn't enough to produce wild-type phenotypes. Recessive wheaten might produce 1/4 signal or homozygous ey ey would produce 1/2 signal and heterozygous ey e+ would produce 3/4 (1/4 + 1/2)signal. The ey ey homozygotes would not produce enough signal and be wheaten, but the ey e+ heterozygotes do.
E and ER would be dominant to wheaten because the E and ER gene products produce enough signal in the heterozygotes to continue to produce the black phenotypes.
The E allele would be expected to send the most signal and has been found to not be regulated properly and seems to always send signal whether the receptor binds hormone or not. So it is always on and always sending signal to make black pigment. It has two mutations that in mammals are associated with black fur. One of the mutations is found in black wooled sheep, and the other is found in black furred mice. The E allele combines those two mutations. The probable birchin allele has only one of these (the one found in mice) two mutations and so may not send as much signal as the E allele product. The Fayoumi birchin allele has a totally different mutation that has not been found in mammals at this time, but produces a similar phenotype to the birchin (ER) allele.
The funky allele that I found in Buff Rocks has the dominant wheaten mutation and the Fayoumi birchin mutation combined. The two mutations should do different things to the gene. Beats me what it looks like without the columbian restrictors.
------------------------------------------
In summary, there have been 7 E locus alleles sequenced:
![]()
-----------------------------------------------------------------------------
* Images used in the following photos are from Feathersite, and the OEGBCA website.
These eumelanin enhancing genes are responsible for extending the typical black areas of the wild type in to the pheomelanin areas. The Brassy Back variety is a good example:
They are also responsible for extending eumelanin in the Quail and Vorwerk varieties: both Buff Columbian based varieties:
Obviously there are differing eumelanin enhancing genes/genotypes in the Quail and Vorwerk varieties, as both varieties can be produced with the base genes: eb Brown, Co Columbian plus eumelanin enhancer(s). Eumelanin enhancing genes are also found in E solid black based varieties. The Ml gene is found in the secondary pattern varieties: Spangled & Doubled Laced and Single Laced. -------------------------------------------------------- Melanotic allele: Ml (1971):First described by Moore and Smyth (1971), after extracting it from a randombreeding recessive white population previously described by Jeffrey (1947). Ml is incompletely dominant autosomal mutation that extends eumelanin into the normally red areas of pyle-zoned fowl, while having little effect on the chick down colour. Heterozygotes on wild-type (e+) or brown (eb) backgrounds are distinctly darker, particularly in the hackle and head; however, Ml/ml+ has little effect on eWh/eWh females. Homozygotes approach self blackness, but the salmon pigment of the wildtype and wheaten females remain evident.
Ml (Melanotic) is sex influenced, autosomal incompletely dominant gene. This means that heterozygote hens with only one dose of the black enhancer (Ml/ml+) show very little pheomelanin (gold or silver) on ER (birchen) base (ie acts like a dominant gene). Whereas, heterozygote roosters with only one dose of Ml (Ml/ml+) show more gold on the hackles, wing bows, etc, when ER (i.e, acts like an incompletely dominant gene). Ml homozygous (Ml/Ml) & on its own doesn't cover all pheomelanin areas on e+ & ER male birds (although covers a lot), & doesn't cover salmon areas of e+ (eg, as in Brassy Backs) & eWh hens. Therefore, other eumelanin enhancers are needed to change the gold/silver areas to black (eumelanin). Ml isn't expressed in day-old chick down, unless in combination with other mutations, eg eWh Co Ml produces a heavily melanised chick down - research by Campo & Oronzco - Melanised Prat. Campo & Oronzco (1986) (and Campo, 1991) also noted that two doses of Ml is needed to produce the dark chick down, & quail pattern in adult eWh Co birds. Unsuccessful efforts to determine the genetic basis for certain “recessive black” lines (Punnett, 1957, Smyth, 1976) suggest the existence of more than one eumelanin intensifying mutation. References: Campo, J.L. and Orozco, F. (1986) Genetic basis of the melanotic Prat phenotype. British Poultry Science. 27:361-367. Campo, J.L. (1991) Use of the sex-linked barring (B) gene for chick sexing on an eumelanotic columbian background. Poultry Science. 1991 Jul;70(7):1469-73. Jeffrey, F.P. (1947). Plumage color genes in White Plymouth Rocks and White Wyandottes. Poultry Sci. 26:526-528. Moore, J.W. and Smyth, J.R. (1971). Melanotic, key to a phenotypic enigma of the fowl. J.Hered. 62:214-219. ------------------------------------------------------ Crawford (1986)- eumelanising gene 0.3 cM to Peacomb locus:Crawford segregated a eumelanising gene, closely linked with the P (pea comb) locus, only 0.3 map units apart (from "Poultry Breeding & Genetics, 1990). Carefoot (1990)- Linkages of Ml with Peacomb locus:Carefoot, 1990 tested linkages again of Ml (Melanotic) with P (peacomb) loci, and found the loci to be 46 map units apart. It was suggested from this evidence that two eumelanising genes were in existance, ie Crawford's eumelanising gene was different to Ml - Melanotic. Therefore, Charcoal (cha) was suggested by Dr Crawford for the recessive gene linked with P by 0.3% (Carefoot, 1990). Smyth (1994)- Charcoal:Smyth listed the Charcoal gene with the dominant symbol - Cha (Smyth, 1994). Carefoot (2001)- Black Hackle of the Indian Game:Carefoot crossed Indian Games (pea-comed, black hackled) with Barnevelders (single-combed, partially black striped hackled), to determine the genetics of the neck hackle eumelanising gene found in Indian Games. He found the black hackled eumelanising gene to be closely linked with the P (pea comb) locus, and to be a dominant gene. Therefore, Carefoot suggested the eumelaniser appearred to be allelic to, if not a further manifestation of the Charcoal gene. *note: Carefoot mentioned in the above report that Smyth (pers. com.) investigated in Brassy Back OEGB a potentially new eumelanin extension that had a dominant charcoal effect on wild type. References: Carefoot, W.C. (2001) Inheritance of the Black Hackle of the Indian Game bantam. British Poultry Science. 2001 May;42(2):274-5. Crawford, R.D. (1986) Linkage between pea comb and melanotic plumage loci in chickens. Poultry Science 65:1859-1862. Smyth, J.R. Jr. (1994) Melanin Pigmentation: Its Biological Roles, Inheritance and Expression in the Chicken. University of Massachusetts, Amherst. -------------------------------------------------------- Cote (1976) - Self Black Plumage:Many examples of self black breeds, but similarity is more phenotypic than genotypic. Most are based on E or ER, but not all. Eumelanin enhancing genes found in E & ER based self blacks. Cote (1976) studied nine self black & one birchen male (1976), all carrying eumelanising modifiers. Ml found in only 5 out of 9 (ie not in birchen, Black P.Rock, Blue Frizzle & Black OEG). It was linked with the black lacing gene (ie Pg) except in a Mottled Ancona that was homozygous for Ml & nonlacing (pg+). Other eumelanin intensifiers were also present, even in the birchen, but they were not identified individually. None of the E heterozygotes developed solid black plumage in the absence of eumelanising modifiers. Eight of the nine males also carried eumelanin restrictors, but only 2 of these carried Co. No birds were carrying Db (tested with Db/Db and Co/Co tester lines)(ed. Crawford, 1990). Cote's research birds......
* References to Cote's research: Crawford, R.D (ed.)(1990). Poultry Breeding and Genetics. Elsevier, Amsterdam. Jeffrey, F.P. (1976) Bantam Chickens, ABA . Smyth, J.R. Jr. (1976) Genetic Control of Melanin Pigmentation in the Fowl. University of Massachusetts, Amherst. -------------------------------------------------------- Smyth's eb Massachusetts Recessive Black line:Totally black, breeds true. Dark Brown down (eb) and white undercolour in the adult males. Shanks and beaks are non-black (Smyth 1976). When crossed to e+/e+ and eb/eb, its net effect is “incomplete dominance”. Three and possibly four genes are hypothesised as contributing to the non-E self black plumage (ed. Crawford, 1990). * References: -------------------------------------------------------- Fred Jeffrey - Old English Games & eumelanisers:Jeffrey, 1988 – Brassy Back & Quail OEG are e+e+ , plus autosomal recessive eumelanin enhancer (the Quails also with Co). This eumelanin enhancer darkens the chick down significantly, and produces a purplish-black hue in the down. The Ml gene (on its own) does not change the chick down. Jefrrey produced solid black male birds with this eumelanin enhancer & wheaten, but hens were black with gold lacing. Fred Jeffrey used the term “hypothetical ebonies” for unknown eumelanin enhancers. Other genes have been suggested, eg sg (eg, listed by Dr Hollander on his chicken color genetics chart - but used sg as a replacement for Ml)) & mi, but mi later believed by Smyth not a new gene, but the effects of Db (Smth & Malone, 1979). Db (Dark Brown, Columbian-like restrictor) is another sex-influenced autosomal gene, but this time, acts dominant in ER males (ie, only one dose needed for expression- Db/db+) and recessive in females (2 doses needed for expression). You can see how this could be misinterpreted as a recessive eumelanin enhancer. References: Jeffrey, F.P. (1988) Crow wing and recessive black in Old English Game bantams. ABA Yearbook, Amherst, MA. Jeffrey, F.P. & Richardson, W. (1995) Old English Game Bantams as Bred and Shown in the United States. ABA. Smyth, J.R., Jr and G.W. Malone, (1979). Evidence that plumage color genes mi and Db are identical. Poultry Sci. 58:1108-1109 (Abstract). Washburn, K.W., & J.R. Smyth Jr., (1967). A gene for partial feather achromatosis in the fowl. J.Hered. 58:130-134. --------------------------------------- It can be difficult determining new eumelanin enhancers. Both Melanotic & Charcoal genes were identified as different, by using linkage relationships. They are both on the same chromosome, but different distances to other known loci (eg P) on the same chromosome.
The Massachusetts Recessive Black line (eb based, not E or ER) has 3, or possibly 4 genes contributing to the all black plumage. So there are obviously more eumelanin enhancers that have not been formally described by scientists. Cote found other eumelanin enhancers, other than Ml in his studies (including in a Birchen Modern Game). E black lines usually have eumelanin enhancers also. |
![]()
-----------------------------------------------------------------------------
* Images used in the following photos are from Feathersite, and the OEGBCA website.
All these genes not only extend pheomelanin into typical eumelanin areas (Columbian restrictors), but also change the shade of pheomelanin- dilute or darken the gold at various levels.

Co -Columbian

Db - Dark Brown
The Db gene is unusual in that when homozygous, it is a very good columbian restrictor in males, yet only a partial columbian restrictor in females. The Db/Db male phenotype is black-tailed red (black-tailed silver if S/S), with e+, eb, ER, etc alleles. There is a European breed, Villafranquina (eb/eb Db/Db), which has the Black-tailed Red male & secondary patterned female phenotype. The Db gene has little effect on salmon colouring in e+ & eWh hens, diluting the colour slightly (Smyth) (Dr Okimoto's personal experience is that Db slightly intensifies the salmon shade).
The Db gene (when homozygous) significantly changes chick down pattern/colour, in all E locus alleles, except for eWh. This gene was named after its trait of modifying ER - black down to brown shades (in some genotypes, the brown may be restricted to the head region- i.e. black body unchanged). With e+ and eb, the Db gene modifies pheomelanin shade to very pale gold-brown to cream/white, & modifies dorsal striping pattern on the back & head - widens light stripes on the back, & may reduce/fade "arrow-head" stripe on the head. Juvenile pheomelanin feathers of e+/e+ Db/Db may appear at first quite pale on the body & wings, contrasting with salmon-red shades on the breast (for both males & females). In juveniles, a poor autosomal barring pattern may be observed on the top/wings, this generally fading with adult plumage (unless modifiers are present, eg Pg). The following link is of an e+/e+ Db/? rooster, from day-old through to adulthood (note, this rooster also has other eumelanin modifiers present).
Db is linked to Ml - 12cM (approx 22cM to Pg & 32cM to P), i.e., the linkages in order along the chromosome are P-32-Db-12-Ml-10-Pg.
*Note, heterozygous e+/e+ Co/co+ hens may appear similar in phenotype to e+/e+ Db/Db hens, ie appear similar to Ginger Red phenotype.

Mh - Mahogany
* note, the above Welsumers have characteristics similar to Mh traits, but I don't know if it has been substantiated whether this is due to the Mh gene.

Di - Dilute (& possibly eWh Co Db Mh)
* note, a solid buff/red phenotype may be produced various ways. The above birds may or may not have Di (or Db), although both Mh & Di were extracted from a solid Buff Minorca bird by Brumbaugh & Hollander. This Buff Minorca bird's genotype was determined to be:
eyor ebc/? Di/Di Co/Co Mh/Mh db+/db+, plus possibly Cb.
Wheaten has a cream feather undercolor. e+ and eb have a gray feather undercolor even in the presence of Co. eWhe+ Coco+ heterozygotes have the gray feather undercolor and birds that I have that are eWheb CoCo (I crossed a New Hamp to a Columbian Wyandotte) also have the gray feather undercolor, but it is lighter and looks more silvery than gray. If you find that your birds are eWheb and they have a white or cream feather undercolor they may have melanotic. Birds with melanotic often have a white feather undercolor, but this may be an interaction with some other genes because I don't think that they all have a white undercolor.
![]()
---------------------------------------------------------------
(Pg, Ml, Db, mo, B, BSd, Co, Db, Erminette (Er) )
Pg- Pattern gene, Ml- Melanotic, Co- Columbian, Db - Dark Brown
* Images used in the following photos are from Feathersite, and the OEGBCA website.
* Note, the following are only base genotypes. Both Smyth & Carefoot have indicated that their proposed genotypes of secondary patterned varieties (with Pg, etc) are the BASE genotypes for these varieties. They have noted what genes have segregated from test breeding (including linkage testing), but admit freely that there are other modifiers needed to produce exhibition quality birds.
Photo |
Hen Phenotype |
Genotype |
Added mutation from previous variety |
|---|---|---|---|
|
eb - Brown based - stippling |
eb eb |
eb - Brown |
 |
Concentric Pencilling |
ebeb PgPg |
Pg - pattern gene |
 |
Double Lacing |
ebeb PgPg MlMl |
Ml - Melanotic |
 |
Single Lacing |
ebeb PgPg MlMl CoCo |
Co - Columbian |
|
ER- Birchen based |
ERER |
ER - Birchen |
 |
Autosomal Barring |
ERER PgPg DbDb |
Pg, Db - Dark Brown, |
 |
Spangled |
ERER PgPg DbDb MlMl |
Ml - Melanotic |
 |
Single-laced |
ERER PgPg DbDb MlMl CoCo |
Co - Columbian |
------------------------------------------------------------------
The following table lists the most common secondary pattern genotypes:
* click the images to enlarge
* Images used in the following photos are from Feathersite, the OEGBCA website & Dr Corti's website (Erminette image).
Phenotype |
Pattern |
Genotype |
Breed/Variety |
Comments |
|
|---|---|---|---|---|---|
eb (Brown) based varieties: |
|||||
|
|
eb hens, stippling on back & body. |
eb - plus red enhancer(s) |
Dark Brown Leghorn |
- plus red enhancer(s)
|
 |
|
Concentric Pencilling |
eb Pg |
Partridge Wyandotte Brahmas, Cochin, etc |
-
|
 |
|
Double-laced |
1: eb Pg Ml 2: eWh Pg Ml Cha* 3: ey Pg Ml Cha* |
1: Barnevelder- Dark 2 & 3: Indian Games - Dark |
-both Smyth & Carefoot segregated eWh in Indian Games/ Cornish lines, but other alleles have been noted (ey - Morejohn, plus wildtype & dark brown downs) |
|
|
Buff Columbian |
eb Co |
Buff Columbian Wyandotte, Brahma, etc |
-
|
 |
|
Single Laced |
eb Pg Ml Co |
Gold Laced Wyandotte, Cochin, etc |
-
|
 |
|
Melanised Co- Quail |
eb Ml Co |
Quail Belgian Barbu Bantams |
also: e+ Ml Co, eWh Ml Co, or another eumelanin enhancer |
 |
|
Melanised Co- Vorwerk |
eb Co + eumelanin enhancer(s) |
Vorwerk or Golden Lakenvelder |
? Charcoal
|
|
|
Black Tailed Red male- stippled hen |
eb Db |
Villafranquina |
similar to Ginger Red variety (e+ Db) - photos are Ginger Reds -e+ |
 |
|
Black Tailed Red male- autosomal barred hen |
eb Pg Db |
Gold Pencilled Hamburg |
-
|
 |
|
Black-Tailed Red male- buttercup hen |
ebc or eb Pg Db |
Silician Buttercup |
Recent DNA sequencing has indicated that this variety may not be ebc, but eb with other modifiers |
 |
|
Black Tailed Red |
1: eWh Db Co ? 2: ey Co Mh 3: eWh Co Mh |
Black Tailed Red 1: Old English Game 2: Rhode Island Red 3: New Hampshire |
This variety can be developed without Db - eg Rhode.I.Reds - red enhancers |
 |
|
Mottled Buff Columbian |
eb Co mo |
Millefleur Belgian Barbu Bantams, Dutch Booted, Pekins, Speckled Sussex |
-may also be eWh , e+ based
|
ER Birchen based varieties: |
|||||
|
|
Brown Red |
ER |
Brown Red Marans |
-not necessarily with gold breast lacing, as the genetics of the lacing is not known |
 |
|
Black Tailed Red male- autosomal barred hen |
ER Fayoumi Pg Db * Smyth believed these had Co. * Carefoot found no Co in his stock (ie co+) . |
Fayoumi |
- note, the photos are of sex-linked silver versions, not gold. |
 |
|
Autosomal barred, both genders |
ER Pg Db Hf |
Campine |
- henny-feathered
|
 |
|
Gold Spangled- both genders |
ER Pg Ml Db |
Gold Spangled Hamburg |
-sometimes henny feathered - Hf
|
 |
|
Single Laced -both genders |
ER Pg Ml Db Co Hf |
Gold Laced Sebright -possibly Polish Laced varieties |
- henny feathered
|
 |
|
Black Mottled |
ER mo + eumelanin extenders (eg Ml) |
Black Mottled Ancona |
- may also be E - Extended Black based |
 |
|
Laced Blue |
ER Bl/bl+ Pg Ml (Co) + eumelanin extenders |
Blue Andalusion |
- may also be E - Extended Black based -Carefoot found Co, but Campo et. al. found co+ |
 |
Non-laced Blue |
ER Bl/bl+ + eumelanin extenders (eg Ml) |
UK Blue Leghorn, UK Blue Wyandotte, Spanish Blue Andalusian, etc. |
- may also be E - Extended Black based *note, this is not lav - Lavender |
|
 |
|
Sex-linked Dilution |
ER BSd + eumelanin extenders (eg Ml) |
Californa Grey |
- may also be E - homozygous BSd/BSd males are white. |
 |
|
Sex-linked Barred |
ER B + eumelanin extenders (eg Ml) |
Cuckoo |
- may also be E - Extended Black based
|
e+ wild type based varieties: |
|||||
 |
|
Barred
wild type |
e+ B |
Crele Old English Game |
-
|
 |
|
Mottled wild type |
e+ mo |
Spangled Old English Game, Mottled Brown Leghorn, etc |
- Spangled also with pheomelanin enhancers
|
 |
|
Melanised wild type |
e+ Ml |
Brassy Back Old English Game |
- may also be other eumelanin enhancers, eg Cha, etc. |
Miscellaneous varieties: |
|||||
|
|
Buff |
eWh Co Mh Di |
Buff Orpington Buff Minorca |
-may also be other E locus alleles, plus Db, Cb. |
 |
Erminette |
E Er |
Erminette
|
-Incomplete Dominant.
|
|
Moore & Smyth (1972) found that by itself the pattern gene caused an incomplete lace – restricted to the distal tip of the feather. Melanotic (Ml) was required to complete the lace, with the two genes functioning dose dependently. The following diagram is based on a drawing from the following article by Smyth: Genetic Control of Melanin Pigmentation in the Fowl (1976)- available for download at Poultry Science.
This is what they segregated from test breeding Silver Laced Wyandottes (from memory, to an eb test line?): The following are eb Co based:
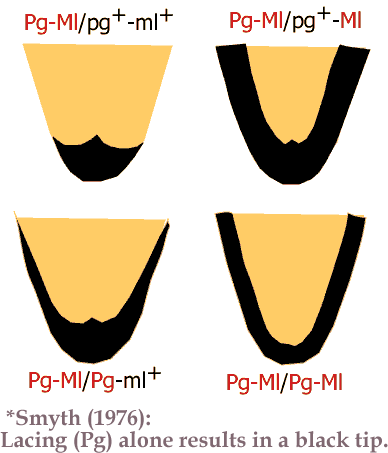
* Note, I’ve changed Lg (from the original drawing) to Pg (& I’ve changed ground colour from silver to gold).
So heterozygous Pg-Ml/pg+-ml+ on Co looks similar to spangling, & Pg/Pg on Co produces an incomplete lace on the tip of the feather.
![]()
----------------------------------------------------------
These genes alter the eumelanin- black areas of the wild type. Some genes change both eumelanin & pheomelanin pigment (eg Lavender, Recessive White, etc).
* Images used in the following table are from Feathersite, and the OEGBCA website, except the Smoky image (from Dr Ron Okimoto), Lavender OEGB image from Al Zwick, photo also found at Yahoo Poultry Genetics website, the Pink-eye Dilute supplied by Kevin Stalder (Stalder Exotics) at the following website, and the sex-linked Chocolate image ( breeder: Dr WC Carefoot, photo supplied by Dr Corti, at the following website).
Photo |
Name |
Gene |
Inheritance Mode |
Comments |
 |
Blue/Splash | Bl | Autosomal Incomplete Dominant | Heterozygous- Blue Homozygous- Splash |
 |
Lavender | lav | Autosomal Recessive | Also dilutes pheomelanin |
 |
Pink-eye Dilute | pk | Autosomal Recessive | Eyes pink-red as chicks. Doesn't appear to dilute pheomelanin (photo of Silver - S bird) |
 |
Dun | ID | Autosomal Incomplete Dominant | One dose darker than two doses. Allelic to I |
 |
Dominant White | I | Autosomal Incomplete Dominant | Fades pheomelanin when homozygous , but not with one dose. |
 |
Smoky | IS | Autosomal Both Dominant & Recessive |
Changes eumelanin - black to smoky shade. One dose visible in chick down- bluish, but two doses needed for adult plumage expression. Allelic to I (Dominant to I). |
 |
Sex-linked Chocolate | choc | Sex-Linked, recessive | Changes black- eumelanin to a chocolate-brown shade |
 |
Recessive White | c | Autosomal recessive | Changes both pheomelanin & eumelanin to white |
Recessive White (c) relates to Tyrosinase in chickens.
There are a few alleles on the c locus, eg c, cre, ca
The cre have very dark red eyes, usually only noticable by shining light into the eyes. The typical recessive whites - c have dark eyes.
There is also a sex-linked imperfect albinism (sal) mutation, proposed to be on the S (silver) locus. This one changes eye pigment to pinkish- through to red. E birds, both down & adult plumage, are changed to "light greyish tan", while red columbian birds -down is changed to "creamy", with adults "tinge with red". Order of dominance on the S locus is: S, s+, sal, i.e. sal is recessive to the wild type sex-linked gold gene. Silversides & Crawford (1991) also noted another allele, a mutation that occurred at the University of Saskatchewan , i.e sex-linked imperfect albinism: Sal-s
Both ca and sal albinos have health associated problems (eg, eyesight problems, higher embryo mortality rates, etc).
References:
Crawford, R.D (ed.)(1990). Poultry Breeding and Genetics. Elsevier, Amsterdam.
Silversides F.G, Crawford R.D. (1991). Phenotypic, embryonic, and neonatal effects of a gene for sex-linked imperfect albinism (Sal-s) in chickens. Poult Sci. 1991 Jun;70(6):1306-13.
An autosomal recessive gene. Eye colour appears pink and the plumage colour appeared blue in the birds studied by Warren (1940). The mutation was first found in White Plymouth Rocks by Warren, and later found in another flock of White Plymouth Rocks by Roberts et. al., in 1952. The mutation doesn't appear to modify pheomelanin.
References:
Crawford, R.D (ed.)(1990). Poultry Breeding and Genetics. Elsevier, Amsterdam.
Hutt, F (1949) Genetics of the Fowl" .
The following Pink-eye (pk) are from Stalder Exotics -the mutation originally derived from Dr Hollander. See the full story on these at the following website: http://stalderexotics.com/chicken_genetics.htm
The following description from "Poultry Breeding & Genetics".
"Grey: - autosomal recessive, dilutes black to light to dark brownish-grey. In older birds, the colour bleached to a dirty yellow."
Reference:
Erasmus, J.E., & Liebenberg, C.R. (1958). A New Mutant in Feather Colour of the South African Black Australorp. S. Afr. J. Agric. Sci. 1:227-230.
The mutation occurred in South Africa.
-------------------------------------------------------
There are at least two different genes that give a chocolate phenotype, ie Dun (ID) and Chocolate (choc). Here is a little history on Carefoot's Chocolate Orpingtons.. * translated from Italian -Dr Corti's website Vol 3 -V 12. Cioccolato quote: ----------------------------------------------------------- The change chocolate in the chicken was observed for the first time in 1994, and precisely from Clive Carefoot in its breeding - - -. In 1996 Carefoot it confided an its large disappointment: although had wanted to put itself repeatedly in contact with an Australian breeder than perhaps had observant in the chicken the same change, not ever some ricevette reply. The disappointment was had to to the fact that Carefoot did not want absolutely to attribute itself the fatherhood of a verification already done from other. ----------------------------------------------------------- So the chocolate gene is a relatively new mutation, first observed by Carefoot in 1994. As the chocolate gene is sex-linked recessive, it will breed true to type. Yet the Dun gene (ID) is similar to Blue (Bl), ie incompletely dominant, where heterozygotes (ID/i+) produce a darker Dun colour, yet the homozygotes (ID/ID) produce a much paler Dun colour. The Dun genetics are explained at MSN Polish Chickens website: The difference in Dun expression are shown in the OEGB varieties: OEGB: Fawn (Dun homozygote ID/ID) Fawn Silver Duckwing (Dun heterozygote ID/i+) Dan has written an article at S.P.P.A, explaining the differences between Dun & Blue: http://groups.msn.com/SPPA/dunvsblue101.msnw Dun seems to have variation in expression, from bluish-dun to a hint of chocolate in others. The following Fawn Breasted Red OEGB appears more blue than chocolate, yet another example of heterozygous ID. Fawn Breasted Red OEGB (Heterozygous ID/i+) * Dr Okimoto has mentioned in the past that red enhancers may modify Dun expression. The Dun gene is an allele of the Dominant White (I) locus. The Dun gene (ID) is found in various breeds in the US, but unfortunately given several different variety names, eg "Fawn" in OEGB, "Chocolate & Khaki" in Polish, "Dun" in Wyandottes, etc. So the reference to "Chocolate" in Dun Polish, etc may cause some confusion in the future, if/once the sex-linked Chocolate (choc) stock becomes available for distribution (I'm not aware if Dr Carefoot has released stock to date). Going by the photos I've seen of Chocolate Orpingtons & Dun birds, the Chocolates (choc) seem to have a glossy sheen to them & a richer chocolate colour (no slate-dun shades, as found in ID). * P.S. I'm not aware of what chocolate-like birds in Australia that were referred to by Carefoot. There are brown breasted birds, eg "Brown Breasted Brown Reds", "Brown Breasted Birchen Duckwings" etc, that are a dull mustard brown shade. But I haven't been able to find out their genetics. It doesn't appear like the colour of Carefoot's Chocolate Orpingtons. I've also seen brown laced gold birds in Australian Pit Game Bantams, I imagine the same brown breasted gene(s) involved. The following is a Brown-breasted Brown Red (?) Australian Pit Game Bantam:

![]()
----------------------------------------------------------
* Images used in the following examples are from Feathersite, and the OEGBCA website.
The following groups influence the ground colour (pheomelanin), at various levels:
Eumelanin Diluters:



* Left: Black, Middle: Blue- Bl, Right: Lavender- lav
I've noticed the blue gene seems to dilute the pheomelanin ground colour some, but not to the extreme of homozygous Dominant White or Lavender. Heterozygous Dominant White (I/i+) doesn't change pheomelanin, but homozygous Dominant White fades the expression of pheomelanin. Lavender (lav/lav) dilutes the expression of pheomelanin dramatically to a straw-cream shade.
Columbian Restrictors :

One other point is the influence of columbian restrictors on gold. The Columbian gene lightens the red of the wild type (eg Black Red- partridge, or Black Breasted Red) to an orangy colour. Yet the Dark Brown gene tends to keep the pheomelanin a richer burnt orange-red colour.
-------------------------------------------------
Mh (Mahogany) characteristics;



Dk, Dk1 characteristics (proposed by Kimball, 1955, but not substantiated):





-------------------------------------------------
Sex-linked Silver:


* Sex-linked Silver on e+ wild type
The Sex-linked Silver gene (S) changes most of the pheomelanin (gold-red) of the wild type phenotype to silver-white, except for the salmon breast of the hens. This gene is incompletely dominant in the males.
Di (Dilute) characteristics;


*note, the above pheonotype is an accumulation of columbian-like restrictors (eg, Co, & other possibilities such as Di, etc)

* The British Poultry Standards of the Cream Legbar chicken down (says to refer to the Silver Legbars for description), suggests the down is diluted dramatically. So it would be interesting to read Punnett's & Taylor's research, to see if a description is given for ig chick down.
*A gold diluted phenotype in my d'Uccles, looks very similar to the cream (ig) phenotype in European d'Uccles (ie, Lemon/Citron Millefleur), although the roosters have adult plumage similar shade of gold to the hens (ig males are lighter silvery/gold phenotype to the gold females). But in my d'Uccles the chick down (pheomelanin only) is diluted dramatically (appearing very similar to silver).

The first feathers appear silvery/very light-gold but darken into patches of gold (see below) at about 3 months on - and into a more even shade of lemon (similar to European Citron Millefleur) at adulthood.
The gold diluter has no effect on id+ slate/blue leg colour, and appears to be a recessive gene.
![]()
----------------------------------------------------------
|
----------------------------------------------------------
\\// Home \\//Genetics Theory \\// Chicken Mutations\\// Chromosome Linkages\\// Chicken Variety Genotypes\\//
\\//Genetics Calculators\\// References/Links\\// About\\//






{kind=link}